
|
||||||||||
|
||||||||||
| Favorite Tweets by @thaibirding | ||||||||||
|
||||||||||
 Buy Birds of Thailand on Amazon.co.uk |
||||||||||
| Site Map ; Contributors |
| A
Comparison of Spring Migration Phenology of Bee-eaters and Oriental
Honey-buzzards Pernis Ptilorhynchus at Tanjung Tuan, Malaysia,
2000-01 By Robert DeCandido, Deborah Allen, Reuven Yosef & K. L. Bildstein |
|||
| Tweet | |||
| Note: This article was originally published in the Ardea 92(2), 2004 and was kindly submitted by Robert DeCandido. | |||
| ABSTRACT DeCandido R., D Allen, R. Yosef & K. Bildstein 2004. A comparison of spring migration phenology of bee-eaters and Oriental Honey-buzzards Pernis ptilorhyncus at Tanjung Tuan, Malaysia, 2000-01. Ardea 92(2): 169- 174 Counts of migrating Blue-tailed Bee-eaters Merops philippinus and Bluethroated Bee-eaters M. viridis, as well as five raptor species were made in March 2000 and March 2001 at Tanjung Tuan, a coastal migration watchsite in western Malaysia. Totals of 2226 bee-eaters (12.9 bee-eaters h-1) and 11 441 raptors (66.5 raptors h-1) were counted. The Blue-tailed Bee-eater comprised 60.8% of Merops individuals (1353 birds) that could be identified. The Blue-throated Bee-eater was much less common, comprising 10.0% of the flight (222 counted). Oriental Honey-buzzards Pernis ptilorhyncus comprised 93.1% (10 648 individuals) of the raptor flight. Bee-eater and Honey-buzzard migration peaked during 11:00-16:00h, and both generally preferred westerly winds to make landfall at Tanjung Tuan. Key Words: Merops philippinus - M. viridi - Pernis ptilorhyncus -migration - Malaysia - East Asian Flyway |
|||
| INTRODUCTION Since the 1950s, Oriental Honey-buzzards Pernis ptilorhyncus have been known to migrate northeast from Sumatra across the Straits of Malacca to the west coast of Malaysia each spring (Oakeley 1955; White 1961; Medway & Nisbet 1965; Medway & Wells 1976; Wells 1990). Recently it was discovered that significant numbers of Bluetailed Bee-eaters Merops philippinus and Bluethroated Bee-eaters M. viridis also use the same route each year (Medway & Wells 1976; Fry et al. 1992; Wells 1999). However, information regarding routes, timing and destinations of diurnal migrants on this East Asian Flyway in both spring and fall are not well known (see McClure 1974; McClure 1998; DeCandido et al. 2004). In Southeast Asia, migrating bee-eaters and Honey-buzzards travel in flocks, interspersing flappingflight with soaring in thermals whenever possible. In this paper we analyze the diurnal rhythm of migrating bee-eaters and Honey-buzzards where both make landfall on the Asian mainland after an overwater crossing of at least 38 km. Port Dickson (2°24’N, 101°51’E, sea-level) is a small town on the west coast of Malaysia situated on the Straits of Malacca, approximately 94 km southwest from Kuala Lumpur (Fig. 1-2). It is located at the southern end of a south to north range of mountains that presumably funnels spring migrants north along the western lowlands of the Malay Peninsula (Medway & Nisbet 1965; Wells 1999). The watchsite, known locally as Tanjung Tuan, is situated on the deck of a lighthouse, south of Port Dickson at km 16, approximately 3 km west of the coastal highway. The lighthouse, built by the Dutch in the 18th century, sits atop a rocky peninsula providing a 180- degree view to the west, south and north; it is surrounded by coastal evergreen rainforest in a small, forest reserve. On clear days it is possible to see Sumatra approximately 38 km across the Straits of Malacca to the southwest. This is the shortest overwater distance along the Straits from Tanjung Medang on the island of Pulau Rupat of northwestern Sumatra (Zalles & Bildstein 2000). |
|||
|
|||
| METHODS Migrants were counted during 8-22 March 2000 (104 observation h) and 2-12 March 2001 (68 h) by DA and RDC using 8.5x and 10x binoculars. Observations typically began at 09:00-10:00 h local time and usually ended by 17:00 h. DeCandido identified, counted, and recorded the numbers of bee-eaters and raptors seen, while Allen photographed the migration. Most bee-eaters (75.9%) and almost all raptors (> 99%) were identified to species level. Weather conditions typically were hazy-cloudy with little windin the morning till 10:00 h, then becoming clear but humid. On many days at approximately 10:45 h, a 3-12 km h-1 sea breeze from the west/northwest would begin. On certain days the wind direction and speed changed significantly in subsequent hours. Wind direction was measured by recording the reading shown on the weather vane mounted atop the lighthouse and a handheld compass. We scanned mainly to the west across the Straits of Malacca in the direction of Sumatra from the patio of the lighthouse. Bee-eaters and raptors were considered migrants if they passed west-to-east across an imaginary north-south line and continued west and out of sight past the lighthouse and nearby hills. Occasionally, migrants did not pass the immediate area of the watchsite but continued on a northeastern course over the Straits of Malacca, presumably making landfall north of the lighthouse towards Port Dickson. We pooled the number counted of both Merops species along with all bee-eaters we could not identify to species level. |
|||
| RESULTS In 2000-01, a total of 2226 bee-eaters of two species (12.9 birds h-1) were counted during 26 observation days. These included 1353 Bluetailed Bee-eaters (60.8%), 222 Blue-throated Bee-eaters (10.0%) and 651 unidentified individuals (29.2%). Also, a total of 11 442 raptors (66.5 birds h-1), were counted in migration. Oriental Honey-buzzard was the most common raptor, making up 93% (10 648 total; 61.9 birds h-1) of the flight in 2000-01. Black Bazas Aviceda leuphotes comprised 5.3% (608 total), and Chinese Goshawk Accipiter soloensis (62), Greyfaced Buzzard Butastur indicus (48) and Japanese Sparrowhawk A. gularis (12), accounted for 1.1% (4.6 birds h-1); 64 raptors could not be identified. The majority of bee-eaters (84.0%) were seen from 11:00-16:00 h (Fig. 3). The highest hourly total occurred between 13:00-14:00 h on 21 March 2000 when 101 Blue-tailed Bee-eaters were counted. Once the first flock of bee-eaters arrived at the Cape, scattered flocks would follow for 4-6 h each day on most days, especially after 10 March. Exceptionally, bee-eater migration would last for seven (17 March 2000) to eight (21 March 2000) consecutive hours on a given day. On peak flight days, the arrival of bee-eaters began in the 09:00-10:00 h (17, 20 and 21 March 2000). Flocks were not usually seen after 16:00 h, and no bee-eaters were recorded migrating after 17:00 h. High daily counts of bee-eaters occurred after 15 March 2000, frequently when winds with a westerly component were moderate to strong (> 15 km h-1) as on 16 March 2000 (169 counted), 17 March 2000 (254) and 19 March 2000 (248). Overall, 30.1% of the bee-eater flight in 2000-01 was counted on these three days of moderate to strong winds. However, the highest day counts in both years occurred on days with light (< 10 km h-1) winds: 11 March 2000 (139 counted) and 21 March 2001 (326). Most (95.0%) migrating Oriental Honey-buzzards were seen from 11:00-16:00 h, with a decided peak between 11:00-12:00 h, and a second, smaller, peak between 14:00-15:00 h and small flocks up until 18:30 h only on days of high counts (2 and 12 March 2001). The highest hourly count in 2000-01 was 841 individuals from 11:00- 12:00 h on 9 March 2001. The majority (96.7%) of Honey-buzzards made landfall at the Cape with light to moderate (< 15 km h-1 and usually < 10 km h-1) winds. Generally, light winds with a westerly component produced the best Oriental Honey-buzzard flights. However, in 2001, of the four days with > 1000 Honey-buzzards counted, three had light winds from the NE to E, and the fourth had light northwestern winds. |
|||
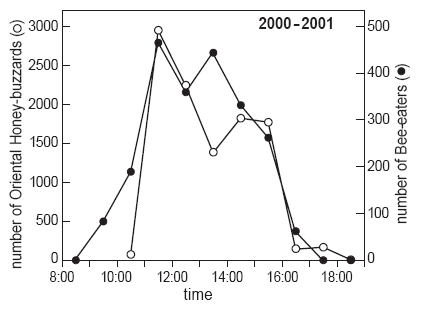 Fig. 3 : Number of bee-eaters and Oriental Honey-buzzards counted in migration by hour of the day at Tanjung Tuan, Malaysia, in March 2000-2001 |
|||
| DISCUSSION Since the 1950s in Malaysia there has been known a significant migration of raptors that are returning each spring from Sumatra to the Asian mainland in the area of Tanjung Tuan. However, only a few published accounts provided information about the daily and seasonal raptor migration phenology at the site (Oakeley 1955; White 1961; Medway and Nisbet 1965; Wells 1990; 1999). Even fewer data exist for the number of individuals of the two Merops species that utilize this same route each spring (see Medway & Wells 1976; Wells 1999). In Southeast Asia, the handful of bee-eater migration reports present data from single days of observation (see David-Beaulieu 1944; David-Beaulieu 1949-50; Melville and Fletcher 1982; Tizard 1996; Evans 2001). The only long-term study of migrating bee-eaters comes from Hong Kong (Carey et al. 2001). Even there, data are based upon a small sample: a maximum 30-year aggregate of 30-40 individuals total, counted by week in spring. At Tanjung Tuan, Oriental Honey-buzzards and bee-eaters showed a diurnal migration pattern typical of soaring birds (Kerlinger 1989), with a late morning peak, an indication of a noon lull and steeply declining passage in the late afternoon. Both species (groups) use thermal soaring and active flight during migration, and readily cross large bodies of open water. In the closely related European Honey-buzzard Pernis apivorus and European Bee-eater Merops apiaster flying speeds were very similar between both species groups (Bruderer & Boldt 2001), but Honey-buzzards showed a wide variation in gliding and flapgliding speeds depending on environmental conditions (Bruderer et al. 1994). The – on average – earlier arrival of bee-eaters at Tanjung Tuan may have been caused by employment of active flight to a greater degree, a lesser dependence on thermals than broad-winged raptors and/or taking advantage of favourable weather conditions in Sumatra (thermals) and Malaysia (onset of seabreeze from the W and NW at about 10:45 h) earlier than Honey-buzzards. Both bee-eaters and Honey-buzzards declined rapidly as migrants after 15:00 h, but why only Honey-buzzards were seen in migration after 17:00 h remains to be determined. Bee-eaters were more likely then Oriental Honey-buzzards to make landfall at Tanjung Tuan when winds were moderate to strong (> 15 km h-1) from the NW-W (DeCandido et al. 2004). Several high daily bee-eater counts occurred on moderate to strong wind days, when simultaneously no Oriental Honey-buzzards were counted in migration. All Honey-buzzard flights at the Cape were correlated with light winds (< 15 km h-1, usually < 10 km h-1). It seems likely that varying numbers of bee-eaters and Honey-buzzards are migrating in all wind speeds and wind directions along a fairly broad front as they approach Tanjung Tuan. Moderate to strong westerly winds concentrated the bee-eater flight at the Cape probably because it was the closest site to make landfall. What factor( s) influence Honey-buzzards to choose Tanjung Tuan to make landfall in large numbers on certain days but not others remain to be determined. Light winds, sometimes with a westerly component, produced the best Honey-buzzard flights, but on other days with the same conditions, few Honey-buzzards appeared. Also, large flights of Honey-buzzards occurred on days with NE-E winds. The influence of wind direction and speed upon migrants at Tanjung Tuan warrants additional investigation, as do the factors that initiate movements of particularly large numbers of bee-eaters and raptors. In any event, conditions in Sumatra probably play the most important role in determining the extent and timing of bee-eater and Honey-buzzard migration at the Cape on any given day. |
|||
| ACKNOWLEDGEMENTS We thank two anonymous reviewers, and especially Rob Bijlsma, for erudite comments and suggestions that greatly improved this manuscript. Anne Arrowsmith designed the two maps. We sincerely appreciate the encouragement and thoughtful advice given by members of the Malaysian Nature Society including Laurence and Audrey Poh, Liew Siew Lan and Ooi Beng Yean, Regina Anthony and Chiu Sein Chiong, Cheang Kum Seng, Lim Kim Chye and Lim Aun Tiah. David Melville and Jevgeni Shergalin, the latter of the Hawk Mountain Sanctuary Acopian Center for Conservation Learning, provided the authors with important reference material regarding Southeast Asian Merops migration. Dr. William Duckworth of the Wildlife Conservation Society suggested many helpful ideas, as well making us aware of the historical information regarding Merops spp. in Indochina. This is publication number 123 of the Hawk Mountain Sanctuary. We dedicate this paper to the memory of Laurence Poh, 1952-2004. |
|||
| REFERENCES Bruderer B., S. Blitzblau & D. Peter. 1994. Migration and flight behaviour of Honey Buzzards Pernis apivorus in southern Israel observed by radar. Ardea 82: 111-122. Bruderer B. & A.Boldt 2001. Flight characteristics of birds: I. radar measurements of speeds. Ibis 143: 178-204. David-Beaulieu A. 1944. Les oiseaux du Tranninh. Université Indochinoise, Hanoi. David-Beaulieu A. 1949–1950. Les oiseaux de la Province de Savannakhet (Bas-Laos). L’Oiseau et R.F.O. 19: 41–84, 153–194; 20: 9–50. Carey G.J., M.L. Chambers, D.A. Diskin, P.R. Kennerly, P.J. Leader, M.R. Leven, R.W. Lewthwaite, D.S. Melville, M. Turnbull & L. Young 2001. The avifauna of Hong Kong. Hong Kong Bird Watching Society, Hong Kong. DeCandido, R., D.A. Allen & R. Yosef. 2004. Merops migration at Tanjung Tuan, Malaysia: an important spring beeeater migration watchsite in South-east Asia. J. Yamashina Inst. Orn. 36: 15-21. DeCandido, R., C. Nualsri, D.A. Allen & K. Bildstein 2004. Autumn 2003 raptor migration at Chumphon, Thailand: a globally significant raptor migration watch site in Southeast-Asia. Forktail 20: 49-54. Duckworth J. W., Davidson, P. & R.J. Timmins 1999. Birds. Pp. 69–159 in Duckworth, J. W.,Salter, R. E. and Khounboline, K. (eds). Wildlife in Lao PDR: 1999 Status Report. IUCN–The World Conservation Union / Wildlife Conservation Society / Centre for Protected Areas and Watershed Management, Vientiane. Evans T. D. 2001. Ornithological records from Savannakhet Province, Lao PDR, January-July 1997. Forktail 17: 21–28. Kerlinger P. 1989. Flight strategies of migrating hawks. University of Chicago Press, Chicago. McClure H.E. 1974. Migration and survival of the birds of Asia. United States Army Medical Component, Southeast Treaty Organization Medical Research Laboratory, Bangkok. McClure, H. E. (1998) Migration and survival of the birds of Asia. Bangkok: White Lotus Press. Medway L. & D.R. Wells 1976. The birds of the Malay Peninsula. Vol. 5. Whitherby, London. Medway L. & I.C.T. Nisbet 1965. Bird Report: 1964. Malayan Nature Journal 19: 160-194. Melville D.S. & W.B. Fletcher 1982. Diurnal observations of bird migration in central and western Thailand. Natural History Bulletin of the Siam Society 30: 49-50. Oakeley R. N. 1955. Migrating hawk-eagles. Malayan Nature Journal 10: 163-64. Tizard R. J. 1996. A preliminary wildlife and habitat survey of the proposed northern extension to the Nakai-Nam Theun National Biodiversity Conservation Area and adjacent Nam Gnouang area, Lao PDR. CPAWM/WCS, Vientiane. Wells D.R. 1990. Malayan bird report: 1982 and 1983. Malayan Nature Journal 43: 116-147. Wells D. R. 1999. The birds of the Thai-Malay Peninsula, Vol. 1. Academic Press, London. White D. H. S. 1961. Migrating Raptors. Malayan Nature Journal 15: 181. |
|||
| Kindly
submitted by: Robert DeCandido, Hawk Mountain Sanctuary, Acopian Center for Conservation Learning, 410 Summer Valley Road, Orwigsburg, Pennsylvania 17961, USA Email: rdcny@earthlink.net Deborah Allen, P.O. Box 1452 Peter Stuyvesant Station, New York 10009 U.S.A. Email: dallenyc@earthlink.net Reuven Yosef, International Bird Research Center, P.O. Box 774, 88106 Eilat, Israel. E-mail: ryosef@eilatcity.co.il Keith L. Bildstein, The Linnaean Society of New York, P.O. Box 1452 Peter Stuyvesant Station, New York, New York 10009 USA. Email: bildstein@hawkmtn.org |
|||
| Tweet | |||

A Guide to Birdwatching in Thailand. Copyright © 2004-2017 thaibirding.com. All rights reserved.